The body-clock is a cluster of neurons connected to the eyes that dictates the rhythm of most bodily functions.
For example, the body-clock, technically known as the Suprachiasmatic Nucleus (SCN), regulates the release of a hormone that determines when we wake up and when we feel sleepy. Research into the function of the body-clock is important: the 2017 Nobel prize for physiology or medicine was awarded to three scientists (Jeffrey Hall, Michael Rosbash and Michael Young) for their significant contributions to the understanding of the inner workings of the body-clock.
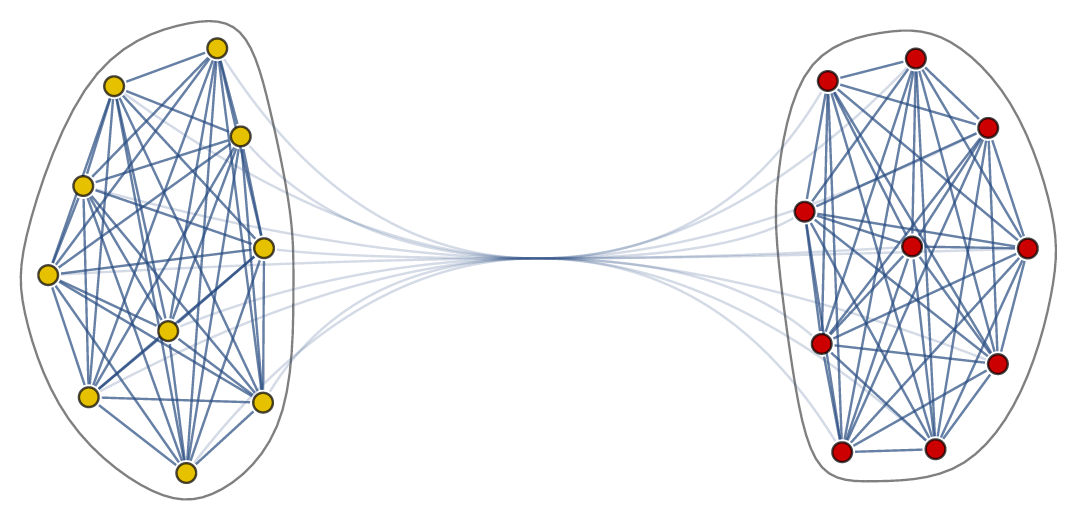
The body-clock, which is a cluster of neurons in the brain, has the same structure in all mammals, which is remarkable. It consists of two groups: two-communities of neurons that are strongly linked within each community and less strongly linked between the communities.
An abstract network representation of two communities of neurons
The neurons "fire" in a periodic way. This means that they pass through a variety of states, culminating in the release of electricity before the process starts again. In order for the body-clock to function optimally, the firing of the neurons needs to be synchronized to some degree. This is achieved because each neuron reacts to the neurons firing around it, by adjusting the moment at which it fires itself. Many experiments have been done to understand the structure and functioning of the SCN better.
Experimental approach
Many experiments have been conducted to understand the influence of the light-dark cycle of the sun on the body-clock. These typically involve rats or mice that are exposed to a regular light-dark cycle, before being put in a changing environment. The goal is to see how the behavior of the animals changes as a result of this change. The activity of the animals is used as a measure of the synchronization of the body-clock. How does the experiment work? The day is divided into time slots and the animal is provided with a running wheel, or something of that type, which measures when the animal is active (running in its wheel). Each time slot in which the animal is active is marked appropriately. When the light-dark cycle is regular, the animal will display a single period of activity, represented by a number of consecutively marked active time slots, followed by a period of inactivity or sleep. Since the body-clock regulates when the animal is active, a regular pattern is a sign of a synchronized body-clock.
After the animal displays a regular pattern, one can change its environment, for example, by exposing it to constant light. This leads to an interesting phenomenon. In some cases the animal will start being active in two periods during the day. Recently, some authors have argued that this is due to the neurons in each community of the body-clock being synchronized within the community but anti-synchronized with the neurons in the other community. This state of the body-clock is called the phase-split state, and the activity pattern observed during the transition to this state can be either smooth or chaotic.
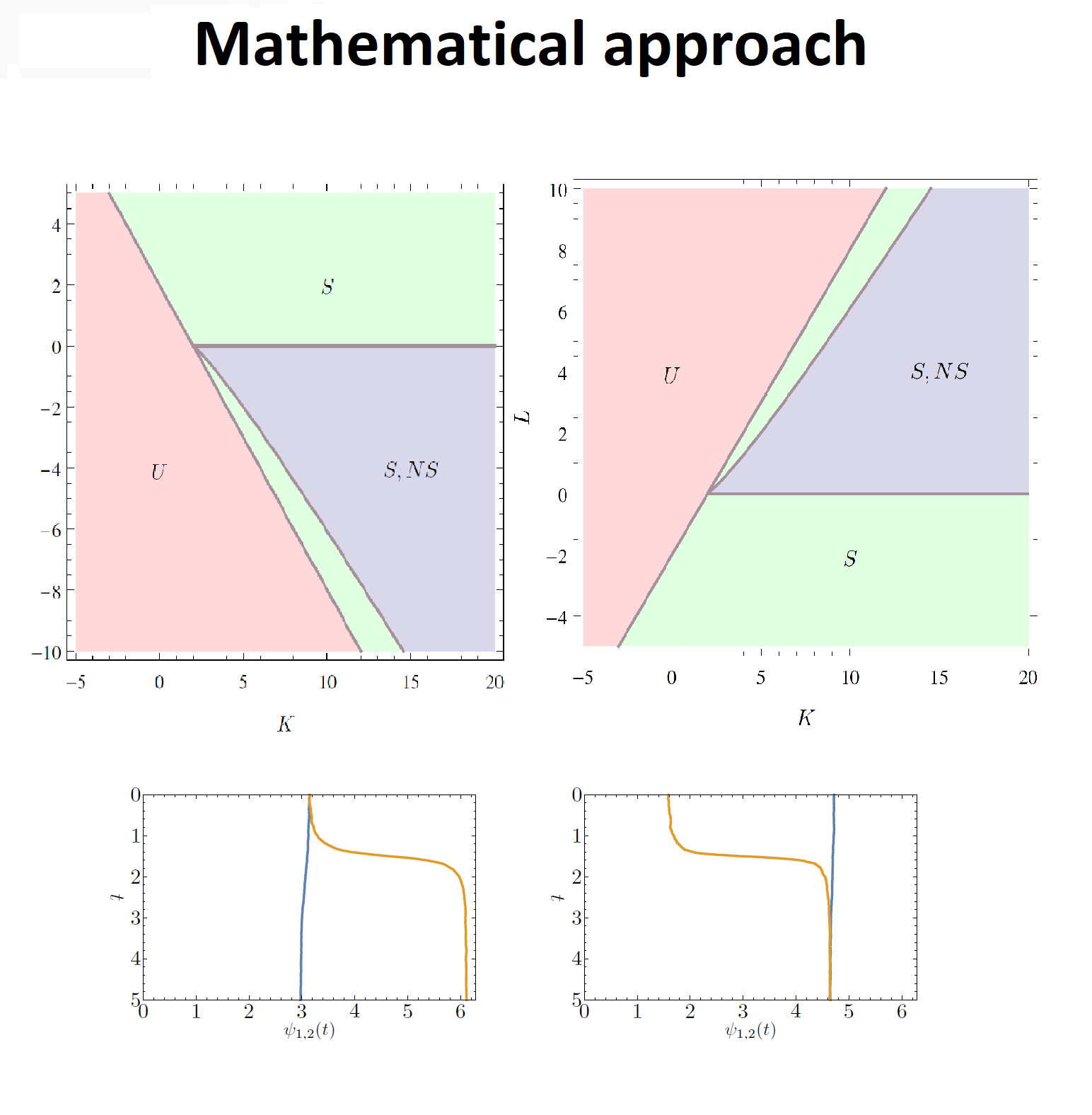
To gain a better understanding of synchronization these experiments are supplemented with mathematical models, which can guide setting up the experiments. One of the mathematical models used to describe synchronization was proposed by Kuramoto in 1975 and this model has been widely studied since then. To reflect the structure of the body-clock, we analyze in this article what happens if we consider the Kuramoto model on a two-community network.
The general idea is the following, we represent the interactions between the neurons in the body-clock as a network, in this network nodes represent thus neurons. In our case the network has two communities, where each community has many edges between the nodes, while there are only few edges between the nodes in different communities. See also the figure above for a visualization of this network.
Below we show how the Kuramoto model on a two-community network works and what it revels about synchrony. We do this in two two self-contained ways: using mathematics and using an interactive animation. Feel free to choose which one you prefer to read first!
Conclusion
The body-clock is a complex network of neurons that is difficult to model mathematically, due to its many intricacies. Above we have seen that we can gain some understanding by using mathematics and simulations, but also that experiments uncovering an interesting phenomenon can play an important role in guiding mathematicians in making choices about which questions are interesting and worthwhile to study.
Specifically, we have found a possible mechanism for the different types of transitions to the phase-split state observed in experiments. Which transition occurs will depend on the interactions strengths, which may be different in different individuals. Much remains to be done in studying the two-community Kuramoto model, and even more when it comes to understanding the body-clock.
The part of the brain that controls the sleeping cycle is made up of two groups of neurons. In order for you to be able to sleep at night, the activity of these neurons needs to be synchronized with the cycle of the sun (e.g. active during the day and inactive at night). When you abruptly change the cycle of the sun (by flying to New York) these two groups need to re-synchronize to the new sun-cycle.
After having a near-fatal accident falling off his horse, Prussian footman Hans Berger (1873—1941) was surprised to learn that his sister had—being kilometers away—him a telegram to let him know that she sensed he was in grave danger.
In times of war, secure communication can be the difference between life and death, or even winning or losing a war. The first to patent a rotor machine in Europe was Arthur Scherbius in 1918. Scherbius’ version of the rotor machine became a commercial success, unlike the other patented machines. Scherbius named his machine Enigma.
The new date for this event is April the 7th 2022 from 19:00 to 21:00 uur. In April the Dutch Academy of Sciences and Arts (KMAW) organizes the "Evening of the Abel prize", a yearly event with as goal to discuss aspects of the research of the latest Abel prize laureates. The event is meant for a […]
Alfred Aho and Jeffrey Ullman received the 2020 A.M Turing Award for their contributions to computer science. Let’s explore how their contributions have shaped the principles, techniques and tools for writing compilers.